Page 18 Volume 11, Número 2, 2003
P. 18
Avelino A, et. al.: A anandamida, um agonista endógeno do receptor vanilóide
Figura 4. Células reactivas com Fos no segmento
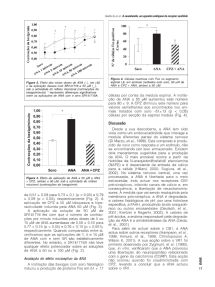
Figura 2. Efeito das várias doses de ANA (_), em µM, espinal L6, em animais instilados com soro, 50 µM de
e da aplicação dessas com SR141716 a 50 µM (_), ANA e CPZ + ANA, ambos a 50 µM.
sob a actividade do reflexo miccional (contracções da
bexiga/minuto); * representa diferenças significativas
entre as aplicações de ANA com e sem SR141716A. células por cortes da medula espinal. A instila-
ção de ANA a 50 µM aumentou este número
para 80 ± 9. A CPZ diminuiu este número para
valores semelhantes aos encontrados nos ani-
mais tratados com soro –51±13 (p < 0,05)
células por secção da espinal medula (Fig. 4).
Discussão
Desde a sua descoberta, a ANA tem sido
vista como um endocanabinóide que interage e
modula diferentes partes do sistema nervoso
(Di Marzo, et al., 1998). Este composto é produ-
zido de novo como resposta a um estímulo, não
se encontrando por isso armazenado. Existem
dois mecanismos sugeridos para a produção
da ANA. O mais provável ocorre a partir da
hidrólise da N-araquidonilfosfatidil etanolamina
(NAPE) e é dependente da entrada de cálcio
para a célula (Hillard, 2000; Sugiura, et al.,
2002). No sistema nervoso central, uma vez
processada, a ANA é libertada para o meio
Figura 3. Efeito da aplicação de ANA a 50 µM e ANA
+ CPZ, ambas a 50 µM, sob a actividade do reflexo extracelular, indo actuar nos receptores CB1
miccional (contracções da bexiga/min). pré-sinápticos, inibindo canais de cálcio e, em
consequência, a libertação de neurotransmis-
sores. À medida que vai sendo readquirida pela
de 0,51 ± 0,09 para 0,73 ± 0,1 (p < 0,05) e 0,79 membrana pós-sináptica, a ANA é degradada,
± 0,09 (p < 0,05), respectivamente (Fig. 2). A a valores fisiológicos de pH, por uma hidrolase
aplicação de CPZ a 50 µM bloqueava a hipe- específica, a FAAH, produzindo ácido araquidó-
ractividade induzida pela ANA 50 µM (Fig. 3). nico ou outros eicosanóides (Deutsch, et al.,
A aplicação da solução de 50 µM de 2001; Kreitzer e Regerhr, 2002). A valores de
SR141716 fez com que o número de contrac- pH ácidos, a enzima responsável pela degrada-
ções por minuto induzidas pelas doses de 5 ou ção da ANA é a amidohidrolase da ANA (Ueda,
10 µM de ANA aumentasse de 0,58 ± 0,10 para et al., 2001).
0,77 ± 0,15 (p < 0,05) e 0,76 ± 0,10 (p < 0,001), Para além de actuar sobre o CB1, a ANA
respectivamente. Quando comparadas entre si, actua sobre outros receptores (Hampson, et al.,
verificamos que as aplicações de 1, 5 e 10 µM 1998; Kimura, et al., 1998; Christopoulos A,
de ANA com e sem SR são estatisticamente Wilson K, 2001). A sua acção sobre o VR1 foi
diferentes. No entanto, o SR141716A não teve primeiro observada por Zygmunt, et al. (1999),
qualquer efeito potenciador sobre as soluções que, in vitro, verificaram que a ANA provocou
de ANA a 50 ou a 100 µM (Fig. 2).
uma libertação do neuropeptídeo relacionado
com o gene da calcitonina (CGRP). Esta acção
Avaliação do efeito nociceptivo da ANA não ocorreu quando foi coadministrada com
A instilação das bexigas com soro fisiológico CPZ, levando a concluir que a ANA actuou DOR
induziu a produção de proteína Fos em 51 ± 17 sobre o VR1. 17